
第十二章 爲何進化論者的主張無效 在前面的章節中,我們根據化石中發現的證據,並從分子生物學的立場審視了進化論的無效性。在本章中,我們將舉出一些被進化論者作爲理論證據提出的生物現象和概念。這些主題尤其重要,因爲它們顯示了沒有支援進化論的科學發現,揭示了進化論者曲解和欺騙的程度。 變異和物種 “變異”是遺傳學中的一個術語,它表示的是引起某種類型或某個物種的個體或群體具有彼此不同的特徵的遺傳現象。例如,地球上的所有人都具有基本相同的遺傳信息,但是有些人眼睛有點斜,有些人有紅發,有些人則有長鼻子,還有一些人個子很矮,所有這些都取決於這種遺傳信息潛在的變異程度。 進化論者把一個物種內的變異作爲進化論的證據。但是,變異並不構成進化的證據,因爲變異只是現有遺傳信息不同組合的結果而已,並沒有將新的特性加入到遺傳信息中。對進化論而言重要的問題是,新的遺傳信息是如何産生新的物種的。
變異自始至終發生在遺傳信息的限制之內。在遺傳學中,這種限制叫做“基因庫”。由於變異,物種的基因庫中的所有特性可能會以各種方式顯示出來。例如,由於變異,尾巴相對較長或腿相對較短的物種可能是爬蟲類,因爲長腿和短腿都可能存在於該物種的基因庫中。然而,往爬蟲類中加入翅膀或羽翼,或者改變它們的新陳代謝,爬蟲類也不會變異成鳥類。這樣的改變需要增加生物體的遺傳信息,而通過變異是行不通的。 達爾文提出他的理論時並沒有考慮到這一點。他認爲不存在變異限制。他在1844年寫的一篇文章中說道:“大部分作者認爲自然界中的確存在變異限制,但是我還沒有發現可以支援這一信條的哪怕一個事實。”143 在《物種起源》中,他引用了不同的變異例子作爲他的理論最重要的證據。 例如,根據達爾文的觀點,動物飼養員將不同品種的牛進行交配,以便産生出一種新的可以産出更多牛奶的品種,並最終轉變成另一個不同的物種。《物種起源》一書中的幾句話最能說明達爾文“無限變異”的觀點: “我能夠看出,熊類完全可以通過自然選擇,獲得更多適應于水中生存的習性,形成愈來愈大的嘴巴,直到成爲鯨魚那樣的龐然大物。”144 達爾文引用如此牽強的例子,是因爲那個時代的人們對科學原始的理解程度。從那以後,在20世紀,根據生物實驗的結果,科學上提出了“遺傳穩定性”(遺傳的自動調節)原理。該原理認爲,試圖通過配種産生新的變異的所有嘗試都沒有結果,因此不同生物物種之間存在著不可逾越的屏障。這意味著動物飼養員絕不可能像達爾文假定的那樣,通過不同品種的牛之間的交配,把牛轉變成另一不同的物種。 駁斥進化論的諾曼·馬克白(Norman Macbeth)在他的《重審達爾文》一書中寫到: “問題的核心在於,生物是否確實毫無限度地發生變化……物種看起來是穩定的。我們所聽到的都是育種員們的失望之辭,他們將工作做到了一定的點上,但看到的只是動植物又回到了它們的起點。儘管經過了兩三個世紀的不懈努力,但他們從來沒能製造出藍玫瑰或黑鬱金香。”145 盧瑟·波班克(Luther Burbank)是史上最傑出的育種家,他針對這方面的事實說道:“存在發育的可能限度,並且這些限度是遵循某種規律的。”146 丹麥科學家W.L.約翰遜(W. L. Johannsen)就此總結道: “達爾文和華萊士強調的變異,不可能有選擇性地超出某個點,即這樣的變異並不包括‘無限偏離’的秘密。”147
同樣地,達爾文在加拉帕戈斯群島(Galapagos Islands)上看到的不同的雀類也只是另一個變異例子,而不是“進化”的證據。近代的觀察表明,這些雀類並沒有經歷過如達爾文所推測的無限變異。而且,大多數被達爾文認爲代表了14種截然不同的物種的雀類,其實是通過相互交配産生的,這就說明它們進行的是同一物種內的變異。科學觀察表明,雀喙其實是“變異”的例子,而它們卻曾被當作所有進化論來源的神話;因此,雀喙不構成進化論的證據。皮特(Peter)和羅斯瑪麗·格蘭特(Rosemary Grant)花費了數年在加拉帕戈斯群島觀察雀的變異,尋找達爾文進化論的證據,但最後不得不得出了這樣一個結論,那就是島上沒有發生過任何“進化”可以促成新遺傳特性的出現。148 抗生素抗性和DDT免疫性不是進化的證據 被進化論者拿來當作進化論證據的一個生物概念,就是細菌對抗生素的抵抗力。許多進化論者把這種抗生素的抗性作爲“生物通過有利突變而發育的例子”。類似的主張同樣應用於昆蟲對殺蟲劑(如DDT)産生的免疫機制。 然而,進化論者在這個問題上同樣錯了。 抗生素是從微生物中産生的、抗擊其他微生物的“分子殺手”。最早的抗生素是由亞歷山大·弗雷明(Alexander Fleming)於1928發現的青黴素,。弗雷明認識到,黴菌中産生可以殺滅葡萄球菌的分子,這一發現被看作是醫學界的一個轉捩點。從微生物中産生的抗生素被用於對付細菌,其結果是成功的。 不久,發現了一些新東西。隨著時間的流逝,細菌形成了對抗生素的免疫力。該機制的原理是這樣的:大量對抗生素敏感的細菌死亡,而另一些不受抗生素影響的細菌快速繁殖並組成了整個細菌隊伍。這樣,所有的細菌就對抗生素免疫了。 進化論者試圖把這種現象看成“細菌適應環境的進化”。 但是,這種膚淺的解釋與真相大相徑庭。在這方面做過詳細研究的以色列生物物理學家、因1997年出版《並非偶然》一書而聞名的李·斯普特納(Lee Spetner),主張細菌的免疫力可以通過兩種不同的機制産生,但沒有任何一種可以做爲進化論的證據。這兩種機制是: 1)轉變抵抗力的基因已存在於細菌中。 2)細菌形成對抗生素的抵抗,是由於突變而失去其基因資料的結果。 斯普特納教授在2001年發表的一篇文章中解釋了第一種機制:
斯普特納進一步說,這不是“進化的證據”: “以這樣的方式産生對抗生素的抵抗力……不是用來說明進化論突變之說的例證。可以證明進化論的遺傳變化,決不能只是向細菌的基因組加入新的資訊,還必須向生物宇宙(biocosm)加入新的資訊。基因水平式的傳遞,僅僅是擴散一些生物中已存在的基因而已。”150
所以,我們在此不能談論進化論,因爲沒有産生新的遺傳信息:已存在的遺傳信息只是在細菌間傳遞。 由突變産生的第二種免疫力機制,也不是進化的例子。斯普特納寫道: “……有時候,微生物通過單一核苷酸的隨機替代作用,獲得對抗生素的抵抗力……鏈黴素由賽爾曼·沃克茨曼(Selman Waksman)和阿爾伯特·斯卡茲(Albert Schatz)發現,並於1944年首次報告;它是使細菌能用這種方式獲得抵抗力的一種抗生素。可是,儘管它們經歷的突變對鏈黴素中的微生物有益,它也不能當作NDT(新達爾文理論)所需的那種突變的例證。抵抗鏈黴素的那種突變在核糖體中表現出來,並使得對抗抗生素分子的分子退化。這種在微生物核糖體表面上的變化,阻止了鏈黴素分子的抗生素作用。這種退化丟失了特性,因此,也丟失了資訊。關鍵在於,不論有多少突變,都不可能從這種突變中獲得(進化)。進化不能建立在積累僅僅使原有特性退化的突變之上。”151 概括地說,與細菌核糖體的突變使細菌抵抗鏈黴素。其原因是突變“分解”了核糖體。也就是說,細菌中沒有增加新的遺傳信息。相反,核糖體的結構被分解了,即細菌“損壞”了。(還有,研究發現,變異細菌的核糖體的功能不如正常細菌的功能。)由於這種“損壞”阻止了抗生素附在核糖體上,所以“抗生素抗性”形成了。 總之,並沒有“形成遺傳信息”的突變的例子。
突變解釋的其他例子,就像上面提到的核糖體的突變,是導致昆蟲“遺傳信息缺失”的現象。 在這種情況下,並不能宣稱細菌和昆蟲的免疫機制就是構成進化論的證據,因爲進化論主張生物通過突變而發育。但是,斯普特納解釋說,無論是抗生素的免疫力還是其他任何的生物現象,都沒有過這種突變的例子:
退化器官謬論
很久以來,“退化器官”的概念作爲進化的“證據”頻繁出現在進化論文獻中。當證明該概念無效後,它終於悄然消失了。但是,一些進化論者仍然相信它,並且有人不時試圖把它作爲重要的進化論證據。 “退化器官”的觀念最早是在一個世紀以前提出的。進化論者宣稱,某些生物的體內存在一些非功能性的器官,這些器官是從祖先那裏遺傳的、不被使用而逐漸退化了的器官。 這個設想很不科學,它的知識根基並不充分。這些“非功能性器官”,實際上是“功能還沒有被發現”的器官。最能說明這種說法的,是進化論者列舉的長長的退化器官表。進化論者S.R.斯卡丁(S.R. Scadding)在《進化理論》(Evolutionary Theory)雜誌上發表的“退化器官能構成進化論的證據嗎?”一文中,承認了這個事實: “既然不可能清楚地鑒別無用的結構,並且所爭論的結構沒有科學依據,那麽我斷定‘退化器官'不能給進化論提供任何特別的證據。”154 德國解剖學家R.威德希姆(R. Wiedersheim)1895年列出的退化器官表中,有闌尾、尾骨等大約100個的器官。隨著科學的發展,人們發現威德希姆表中的所有器官,其實在人體內都有非常重要的作用。例如,被當成“退化器官”的闌尾,其實是人體內防感染的淋巴器官。1997年,這個事實被澄清了:“其他身體器官和組織:胸腺、肝、脾臟、闌尾、骨髓以及一些小的淋巴組織,如咽喉中的扁桃體和小腸中的淋巴集結等,也是淋巴系統的部分。它們同樣幫助人體防止感染。”155
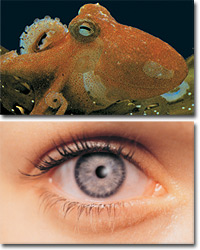
研究亦發現,列入退化器官表中的扁桃體,在保護喉嚨抗感染方面扮演著重要角色,尤其在人的青春期。據發現,位於脊椎骨末端的尾骨支撐著盆骨周圍的骨頭,也是一些較小的肌肉的會聚處,因此,如果沒有尾骨就不可能坐得舒服。在隨後的幾年中,人們認識到,胸腺通過啟動T細胞來啓動人體內的免疫系統;松果腺負責一些重要激素的分泌;甲狀腺對嬰兒和兒童的穩定成長非常有用;腦垂體控制許多激素腺的正常運作。而所有這些器官曾被認爲是“退化器官”。還有,被達爾文看成是“退化器官”的眼睛中的半月襞,實際上負責清潔和潤滑眼球。 進化論者關於退化器官的主張,有一個非常重要的邏輯錯誤。正如我們剛才所看到的,進化論者聲稱的生物的退化器官,都是從它們的祖先那裏遺傳來的。然而,一些所謂“退化了的”器官,並沒有在稱爲人類祖先的生物中發現過!例如,闌尾在被認爲是人類祖先的猿類中並不存在。挑戰退化器官理論的著名生物學家H.伊諾克(H. Enoch),論述了這個邏輯錯誤: “猿有闌尾,與它們沒有直接的親緣關係的較爲低級的猿卻沒有,而一些更爲低級的哺乳動物如負鼠卻又有闌尾。進化論者怎樣解釋這種現象呢?”156 進化論者匆匆提出的退化器官的假想,存在許多邏輯佯謬,並且被證明在科學上是不真實的。人體根本不存在所謂從祖先那裏遺傳來的任何退化器官,因爲人類並非從其他生物偶然進化而來的,而是以現有的完整、完美的形態被創造出來的。 同源神話 生物學中把不同物種之間的結構相似性稱作“同源”。進化論者試圖把這些相似性作爲進化的證據。 達爾文認爲具有相似(同源)器官的生物,彼此之間存在進化關係,並且這些相似器官必定是從共同的祖先那裏繼承來的。根據他的設想,鴿子和老鷹都有羽翼;因此,鴿子、老鷹甚至所有其他有羽翼的鳥類都可以認爲是從共同的祖先進化而來的。 同源是靠不住的論點,它只是憑外觀形體上的相似而提出的假設。自達爾文時代起,至今沒有任何具體的發現來證實這個論點。世界上沒有一個地方發現過假想的同源結構生物的共同祖先的化石。而且,下列發現清楚表明,同源並沒有提供發生進化的證據: 1. 人們發現生物的同源器官屬於完全不同的門,進化論者一直以來不能在它們當中建立任何一種進化關係; 2. 某些具有同源器官的生物的遺傳密碼彼此完全不同。 3. 不同生物同源器官的胚胎發育階段完全不同。 現在我們對這些要點逐一加以分析。 完全不同的生物物種的相似器官 在進化論者不能建立任何進化關係的不同物種之間,有許多同源器官是共有的。翅膀就是其中的一個例子。除了鳥類,作爲哺乳動物的蝙蝠也有翅膀,並且昆蟲甚至一些滅絕的恐龍也有翅膀。進化論者無法在這四種不同種類的動物中,建立任何進化關係或親緣關係。 另一個明顯的例子,就是不同生物的眼睛驚人的相似以及結構上的類似。例如,章魚和人是兩種極其不同的物種,它們之間無法建立任何進化關係,而兩者的眼睛在結構與功能上卻極爲相似。沒有哪個進化論者可以提出一個它們之間的共同祖先,來解釋人和章魚眼睛的相似性。這些以及其他許多例子均表明,進化論者以相似性爲基礎的主張是毫無科學依據的。
實際上,進化論者應爲同源器官說感到尷尬。著名進化論者弗蘭克·索爾茲伯里(Frank Salisbury)談到差異極大的物種具有非常相似的眼睛使同源理論陷入絕境,他說: “像眼睛一樣複雜的器官在其他群體中出現過,如頭足動物、脊椎動物和節肢動物。把這些生物的起源解釋上一次,事情就夠糟糕的了,但根據現代合成理論的觀點把它們的起源想上幾次,我就一頭霧水了。”157
儘管許多生物的形體極爲相似,但這並不支援它們之間有進化關係的主張。胎生動物和有袋動物這兩大哺乳動物種類就是例子。進化論者認爲,這兩種哺乳動物一開始就彼此分開,並完全獨立地進化發育。然而有趣的是,這兩類哺乳動物中有些“配對”幾乎完全相同。美國生物學家迪恩·肯揚(Dean Kenyon)和帕西瓦爾·戴維斯(Percival Davis)評論道: “根據達爾文理論,狼、貓、松鼠、土撥鼠、食蟻獸、鼴鼠和老鼠需要進化兩次:第一次進化成胎生動物,第二次完全獨立地進化成有袋動物。這無異於隨機、不受操控的突變和和自然選擇過程的驚人主張:以某種方式在區別極大的生物體中數次偶然地發展同樣的特徵。”158 進化論生物學家不能把這些極其接近和相似的器官作爲“同源”的例子,這表明從共同的祖先進化的理論是沒有證據的。那麽,對生物中相似結構的科學解釋是什麽?這個問題的答案在達爾文理論統治科學界以前就給出了。科學家卡爾·林奈(Carl Linnaeus)根據生物的相似結構,首次對生物進行了系統化,理查得·歐文(Richard Owen)認爲這些結構是“共同”創造的例子。換言之,相似器官(今天也稱作相似基因)一直以來都是如此,因爲它們是被創造出來用做特殊用途的,而不是因爲它們是從共同的祖先偶然進化而來的。 現代科學發現表明,憑藉相似器官提出的“共同祖先”的主張是不正確的,唯一可能的解釋就是共同創造,這再一次驗證了生物是由真主創造的。 同源在遺傳與胚胎學上的僵局
爲了支援進化論者關於“同源”的主張,不同生物的相似(同源)器官也應該用相似(同源)的DNA密碼來編碼。但是,事情並非如此。相似器官通常由極其不同的遺傳(DNA)密碼控制。而且,不同生物在DNA中相似的遺傳密碼,常常與截然不同的器官相關。 澳大利亞生物化學教授邁克爾·登頓在其《進化論:危機中的理論》一書中,描述了進化論者對同源的解釋在遺傳學上所面臨的尷尬:“同源結構常常列入非同源體遺傳系統中,而同源的概念很少擴展到胚胎學的範圍中。”159 這方面的著名例子,就是在幾乎所有進化論教科書中都引用的四足動物的“五趾骨架結構”。四足動物也就是陸生脊椎動物,它們前後爪子上的指關節都有五趾。儘管它們並不像我們知道的那樣始終看起來有五趾,但由於它們的骨骼結構而被認爲是五趾型的。青蛙、蜥蜴、松鼠或猴子的前後爪也是那樣。鳥類和蝙蝠的骨骼也有這種基本的結構。 進化論者關於所有生物源于一個共同祖先的主張,其長期引用的證據就是五趾型肢。在整個20世紀,生物學方面的所有基礎資源幾乎都引用了這個主張作爲進化強有力的證據。但20世紀80年代的遺傳學發現駁倒了這一進化論主張。人們認識到,不同生物的五趾型肢,是由完全不同的基因控制的。進化論生物學家威廉姆·菲克斯(William Fix)這樣描述這一理論的破滅: “進化論老式的教科書大量使用了同源的概念,指出不同動物的肢骨之間存在明顯的相似性。這樣,人的手臂、鳥的翅膀和鯨的鰭上的”五趾”型肢都表明它們有共同的起源。如果這些形式多樣的結構是由同樣的基因組經過不斷的突變,並以自然選擇的形式轉變,那麽,該理論就有意義了。不幸的是,情況並非如此。人們現在知道,同源器官是通過不同物種中完全不同的基因組合産生的。從同一祖先傳承類似基因的同源概念已經破滅了……”160 還有一點,爲了支援進化論的同源主張,在相似結構的胚胎發育階段,換言之,就是在卵或母體子宮內的發育階段,需要類似的發育,然而在現實中,每種生物的相似結構的胚胎階段是截然不同的。 總而言之,我們可以說,遺傳與胚胎方面的研究已證明,達爾文定義爲“生物從同一祖先進化的證據”的同源概念,根本不能視爲證據。在這方面,科學一再證明了達爾文論點的佯謬。 分子同源性主張的無效性 進化論者把同源性作爲進化的證據,不僅在形態學上是無效的,而且在分子上也是無效的。進化論者認爲,不同生物的DNA密碼或相應的蛋白質結構是相似的,而這種相似性是這些生物來自共同祖先或彼此進化的證據。 然而,分子比較的結果其實根本不支援進化論。表面上看起來非常相似和相關的生物之間存在著巨大的分子差異。例如,對呼吸至關重要的細胞色素C蛋白質,在同類生物中具有難以置信的差異。根據這方面的研究結果,兩種不同爬蟲類之間的差異,要比鳥跟魚或魚跟哺乳動物之間的大。另一研究顯示,同一鳥類間的分子差異,要比這類鳥和哺乳動物之間的差異還大。人們還發現,看上去非常相似的細菌的分子差異,要比哺乳動物和兩棲動物或昆蟲之間的還大。161 在血紅蛋白、肌紅蛋白、激素和基因中也作了這樣的比較,得出的結論也是類似的。162 關於分子生物學領域的這些發現,邁克爾·登頓博士說道:
“生命樹”瓦解 20世紀90年代對生物遺傳密碼的研究,加劇了進化論在這方面進退維谷的尷尬境地。在這些實驗中,沒有採用早些時候局限於對蛋白質序列的比較,而用“核糖體RNA”(rRNA)序列來比較了。從這些發現中,進化論科學家試圖建立一棵“進化樹”。然而,結果使進化論者感到失望。根據法國生物學家赫維·菲利普(Hervé Philippe)和帕特裏克·福特瑞(Patrick Forterre)1999年寫的一篇文章,“隨著可用的序列越來越多,大多數蛋白質的系統發展彼此抵觸,rRNA樹也一樣。”164 除rRNA的比較外,對生物基因中的DNA密碼也進行了比較。但是,結果與進化論者假設的“生命樹”恰恰相反。1999年,分子生物學家詹姆士·A.·萊克(James A. Lake)、拉維·傑因(Ravi Jain)、瑪麗亞·C.·裏維拉(Maria C. Rivera)在一篇文章中對此進行了詳細說明:
對蛋白質的比較和對rRNA或基因的比較,都不能證實進化論的假設。伊利諾斯大學享有聲譽的生物學家卡爾·沃斯(Carl Woese)承認,在分子發現面前,“系統發生論”的概念已失去了它的意義:
分子比較的結果,不是對進化論的贊同而是對它的反對。在1999年《科學》雜誌上發表的“是時候根除生命樹了嗎?”一文中,進化論也承認了這個事實。文章的作者伊麗莎白·彭尼斯(Elizabeth Pennisi)說,進化論生物學家對基因的分析與比較,本來想建立“生命樹”,但結果正好相反,並說“新資料正混亂進化論的畫卷”:
簡而言之,隨著分子生物學的發展,同源體的概念喪失了更多的基礎。對蛋白質、rRNA和基因組成的比較顯示,按照進化論的說法是近親的生物其實彼此間差別迥異。1996年的一次研究種使用了88個蛋白質序列,把兔子歸在靈長目而非嚙齒目動物中;1998年對19種動物的13種基因的分析,把海膽歸到了脊索動物中;1998年基於12種蛋白質實施了另一項研究,得出母牛比起馬類而言更接近鯨類。分子生物學家喬納森·威爾斯(Jonathan Wells)這樣歸納了2000年中的情況: 不同分子引起的(生命)樹的不一致性,以及由某些分子分析導致的奇怪的樹,現在將分子系統發生論推入了危機之中。168 “分子系統發生論”面臨著危機——這意味著進化論同樣面臨著危機。(系統發生論是指所謂的各種不同生物之間的“家族關係”,它是進化論的假想基礎。)科學再一次瓦解了生物是彼此演變而來的論點,顯示了所有生物群體都是獨立創造出來的。 胚胎學重演的神話
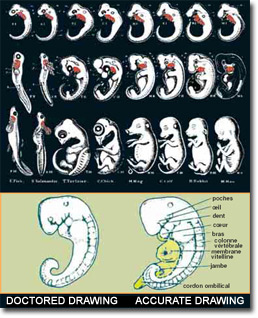
以前所稱的“重演論”早就從科學文獻中除去了,但仍有一些進化論的出版物把它作爲科學事實提出來。“重演”一詞是對進化論生物學家厄恩斯特·赫克爾(Ernst Haeckel)在19世紀末提出的“個體發生學重演族系發展史”斷言的概括。 赫克爾的理論假定,活胚胎再次經歷了其假定祖先經歷的進化過程。他的理論聲稱,人的胚胎在母體子宮裏的發育過程中,首先顯示魚的特徵,然後是爬行動物的特徵,最後才是人的特徵。 據證實,這個理論完全是僞造的。人們現在知道,假定在人類胚胎早期出現的“鰓”,實際上是人的中耳導管、甲狀旁腺和胸腺的初始階段。胚胎被比作“卵黃囊”的部分其實是爲嬰兒造血的育兒袋。被赫克爾和他的追隨者鑒定爲“尾巴”那個部分,實際上是脊椎,它之所以像尾巴,只是因爲它在腿部成形前就已經成形了。
在科學界,這些都是被普遍公認的事實,甚至連進化論者也都接受了這個事實。新達爾文主義的創始人之一喬治·蓋羅德·辛普森(George Gaylord Simpson)寫道:
我們在《美國科學家》雜誌上出版的一篇文章中看到:
“重演”的另一有趣之處是,厄恩斯特·赫克爾本人是個作假高手,他爲了支援自己提出的重演論僞造了圖畫。赫克爾的僞造企圖說明魚和人的胚胎彼此相似。當被曝光後,他唯一的辯護之辭就是其他進化論者也犯過類似的錯誤:
的確有“數百位同道人士,他們中有許多是最值得信賴的觀察者和最值得尊重的生物學家”,他們的研究充滿了偏頗的結論、曲解甚至是僞造。這是因爲他們一向視自己爲進化論的衛道士,儘管不存在任何支援它的科學證據。 |
||||||||||






144. Charles Darwin, The Origin of Species: A Facsimile of the First Edition, Harvard University Press, 1964, p. 184
145. Norman Macbeth, Darwin Retried: An Appeal to Reason, Harvard Common Press, New York : 1971, p. 33
146. Ibid, p. 36
147 Loren Eiseley, The Immense Journey, Vintage Books, 1958. p. 227
148 H. Lisle Gibbs and Peter R. Grant, "Oscillating selection on Darwin's finches", Nature, 327, 1987, pp. 513; For more detailed information, please see Jonathan Wells, Icons of Evolution, 2000, pp. 159-175.
149 Dr. Lee Spetner, "Lee Spetner/Edward Max Dialogue : Continuing an exchange with Dr. Edward E. Max", 2001, http ://www.trueorigin.org/spetner2.ap
150 Ibid.
151 Ibid.
152 Francisco J. Ayala, "The Mechanisms of Evolution", Scientific American, Vol. 239, September 1978, p. 64
153 Dr. Lee Spetner, "Lee Spetner/Edward Max Dialogue: Continuing an exchange with Dr. Edward E. Max", 2001, http ://www.trueorigin.org/spetner2.ap
154 S. R. Scadding, "Do ‘Vestigial Organs’ Provide Evidence for Evolution?", Evolutionary Theory, Vol. 5, May 1981, p. 173
155 The Merck Manual of Medical Information, Home edition, New Jersey : Merck & Co., Inc. The Merck Publishing Group, Rahway, 1997
156 H. Enoch, Creation and Evolution, New York : 1966, pp. 18-19
157 Frank Salisbury, "Doubts About the Modern Synthetic Theory of Evolution", American Biology Teacher, September 1971, p. 338
158 Dean Kenyon & Percival Davis, Of Pandas and People: The Central Question of Biological Origins, Dallas : Haughton Publishing, 1993, p. 33
159 Michael Denton, Evolution: A Theory in Crisis, Londres, Burnett Books, 1985, p. 145
160 William Fix, The Bone Peddlers: Selling Evolution, New York : Macmillan Publishing Co., 1984, p. 189
161 W. R. Bird, The Origin of Species Revisited, Thomas Nelson Co., Nashville : 1991, pp. 98-99 ; Percival Davis, Dean Kenyon, Of Pandas and People, Haughton Publishing Co., 1990, pp. 35-38
162 W. R. Bird, The Origin of Species Revisited, pp. 98-99, 199-202
163 Michael Denton, Evolution: A Theory in Crisis, Londres : Burnett Books, 1985, pp. 290-91
164 Hervé Philippe et Patrick Forterre, "The Rooting of the Universal Tree of Life is Not Reliable", Journal of Molecular Evolution, Vol. 49, 1999, p. 510
165 James Lake, Ravi Jain et Maria Rivera, "Mix and Match in the Tree of Life", Science, Vol. 283, 1999, p. 2027
166 Carl Woese, "The Universel Ancestor", Proceedings of the National Academy of Sciences, USA, 95, (1998) p. 6854
167 Ibid.
168 Jonathan Wells, Icons of Evolution, Regnery Publishing, 2000, p. 51
169 G. G. Simpson, W. Beck, An Introduction to Biology, New York, Harcourt Brace and World, 1965, p. 241
170 Keith S. Thompson, "Ontogeny and Phylogeny Recapitulated", American Scientist, Vol. 76, May/June 1988, p. 273
171 Francis Hitching, The Neck of the Giraffe: Where Darwin Went Wrong, New York : Ticknor and Fields 1982, p. 204